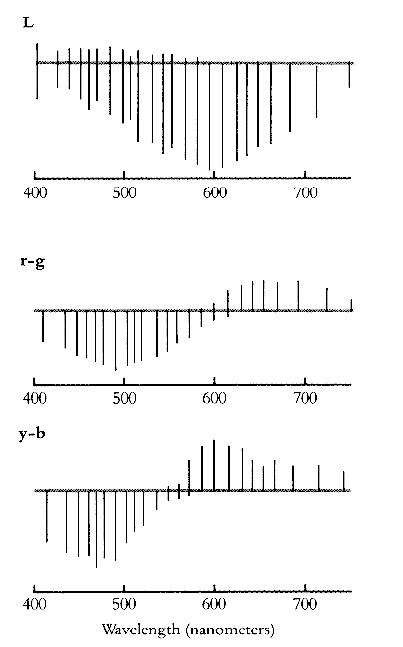
Physiological evidence for Hering's theory came over half a century later. Svaetichin (1956) found in the eyes of teleost fish, by testing them with monochromatic lights in the spectrum of 400 - 700 nm.
De Valois (1958) found in the LGN of macaque monkeys three types of similar cells,
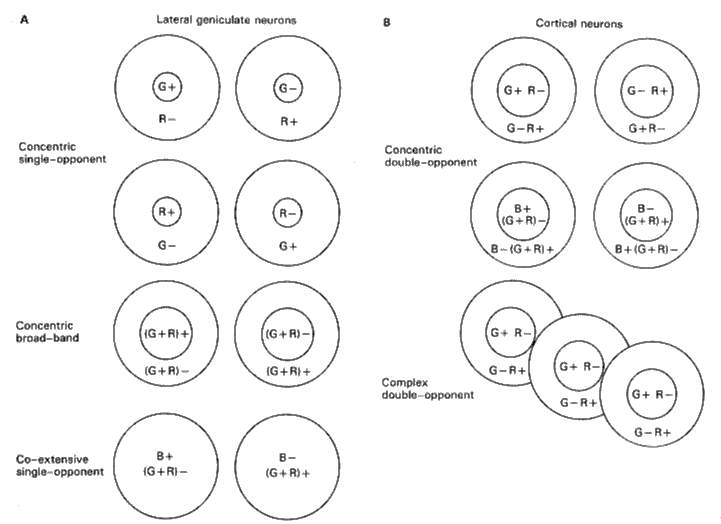
Hering's opponent color theory was further supported Wiesel and Hubel (1966) who used small spots of colored lights and found in the parvocellular layers of LGN (the upper 4 layers) whose receptive fields have antagonistic and color-opponent structures.
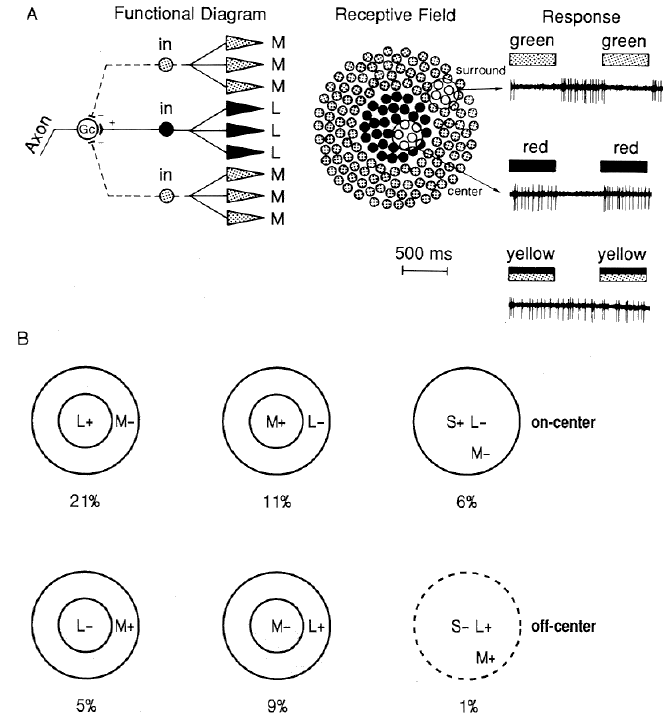
The simplest possible way a r+ center g- surround RF of a type 1 LGN cell could be built is that the RF center gets excitatory input from red cones and the RF surround gets inhibitory input from the green cones. But it is also possible for the entire RF to be fed with both red and green cones with their density spatial distributions as shown in the figure. The overall effect will be an excitatory center for red and an inhibitory surround for green.
Livingstone and Hebel later (1981) discovered double component cells in the blobs of V1. Their RFs are a few times larger than that of the color-opponent cells in LGN.
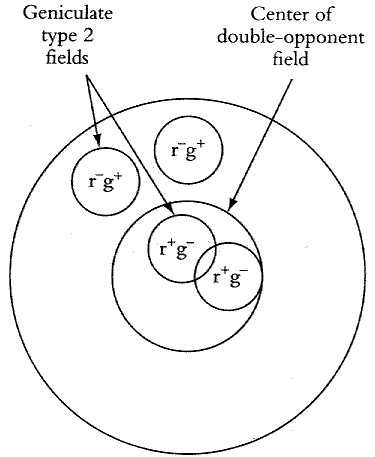
In cortical area V1, the r+g- center r-g+ surround RF of a double opponent could be formed by its connections to the type 2 cells in the LGN. The r+g- center may be fed by mostly the r+g- type 2 LGN cells, while the r-g+ surround may be fed by the r-g+ type 2 LGN cells, as shown in the figure.